Холестерин синтезін реттеу
Холестерин синтезін реттеу - оның негізгі ферменті (HMG-CoA редуктаза) әртүрлі жолдармен жүзеге асырылады.
HMG редуктазасының фосфорлануы / депосфорлануы. Инсулин / глюкагон коэффициентінің жоғарылауымен бұл фермент фосфорилатқа түсіп, белсенді күйге өтеді. Инсулиннің әрекеті 2 фермент арқылы жүзеге асырылады.
HMG-CoA редуктазасы киназды фосфатаза, ол киназаны белсенді емес депосфорланған күйге айналдырады:
Фосфотаза HMG-CoA редуктаза оны депосфорланған белсенді күйге айналдыру арқылы. Бұл реакциялардың нәтижесі HMG-CoA редуктазасының депфосфорланған белсенді формасының түзілуі болып табылады.
Демек, сіңу кезеңінде холестерин жоғарылайды. Осы кезеңде холестерин - ацетил - КоА синтезі үшін бастапқы субстраттың қол жетімділігі артады (құрамында көмірсулар мен майлар бар тағамды жеу нәтижесінде, глюкоза мен май қышқылдарының ыдырауы кезінде ацетил Коа түзіледі).
Постабсорбциялық жағдайда протеингеназа А арқылы глюкагон HMG - CoA - редуктазаның фосфорлануын ынталандырады, оны белсенді емес күйге айналдырады. Бұл әрекет глюкагонның HMG-CoA редуктазасының фосфорлануын және инактивациясын ынталандырады, осылайша HMG-CoA редуктазасын фосфорланған белсенді емес күйде ұстайды. Нәтижесінде постабсорбция кезеңінде және ораза кезінде холестерин синтезі эндогендік синтезге кедергі келтіреді. Егер тамақ құрамындағы холестерин мөлшері 2% -ға жеткізілсе, онда эндогенді холестерин синтезі күрт төмендеді. Бірақ холестерин синтезін толық тоқтату пайда болмайды.
Тағамнан келетін холестериннің әсерінен холестерин биосинтезінің тежелу дәрежесі әр адамға байланысты өзгереді. Бұл холестериннің түзілу процестерінің жеке ерекшелігін көрсетеді. Холестерин синтезінің қарқындылығын төмендету арқылы оның қандағы концентрациясын төмендетуге болады.
Егер бір жағынан холестеринді тамақпен қабылдау және оның ағзадағы синтезі мен екінші жағынан өт қышқылдары мен холестериннің шығуы арасындағы тепе-теңдік бұзылса, тканьдер мен қандағы холестерин концентрациясы өзгереді. Ең ауыр зардаптар қандағы холестерин концентрациясының жоғарылауымен байланысты (гиперхолестеринемия), ал атеросклероз мен холелитиаздың даму ықтималдығы артады.
Отбасылық гиперхолестеринемия (HCS) - бұл форма әлдеқайда жиі кездеседі - 200 адамға шамамен 1 науқас. HCS-тегі мұрагерлік ақау LDL-нің жасушаларға сіңуін бұзады, сондықтан LDL катаболизмі жылдамдығының төмендеуі болып табылады. Нәтижесінде қандағы LDL концентрациясы, сондай-ақ холестерин жоғарылайды, өйткені LDL-де оның мөлшері көп. Сондықтан, HCS-мен холестеринді тіндерге, атап айтқанда теріге (ксантомалар), артерия қабырғаларына тұндыру тән.
HMG-CoA редуктазының синтезін тежеу
Холестериннің метаболизм жолының соңғы өнімі. Ол HMG-CoA редуктаза генінің транскрипция жылдамдығын төмендетеді, осылайша өзінің синтезін тежейді. Бауыр холестериннен өт қышқылдарын белсенді түрде синтездейді, сондықтан өт қышқылдары HMG-CoA редуктаза генінің белсенділігін тежейді. HMG-CoA редуктаза шамамен 3 синтезден кейін болғандықтан, бұл холестерин ферментінің синтезін тежеу тиімді реттеу болып табылады.
Іздегеніңізді таппадыңыз ба? Іздеуді пайдалану:
Холестерол эфирінің алмасуы
Холестерин қорында холестерин мен холестеринді эфирлер бар, олар жасушаларда да, қандағы липопротеидтерде де болады.
II бөлім Метаболизм және энергия
Жасушаларда холестеринді эфирлеу ацил-КоА-холестерин-ацилтрансферазаның (AChAT) әсерінен болады:
Ацил-КоА + холестерин - * HS-KoA + ацилхолестерин
Адам жасушаларында негізінен линолейхолестерин түзіледі. Бос холестериннен айырмашылығы, оның жасуша мембраналарындағы эфирлер өте аз мөлшерде болады және негізінен липидтер тамшыларының бөлігі ретінде цитозолда болады. Эфирлердің түзілуін бір жағынан артық холестеринді мембранадан шығару механизмі, екінші жағынан, жасушада холестеринді сақтау тетігі ретінде қарастыруға болады. Холестерол эфирлерін гидролиздейтін эфираза ферменттерінің қатысуымен ресурстар жұмылдырылады:
Ацилхолестерин + H 2O - * Май қышқылы + Холестерол
Эфирлердің синтезі мен гидролизі көптеген жасушаларда болады, бірақ әсіресе бүйрек үсті безінің клеткаларында белсенді: бұл жасушаларда барлық холестериннің 80% -ына дейін эфирлер қатысады, ал басқа жасушаларда ол әдетте 20% -дан аз болады.
Қанның липонротеиндеріндегі эфирлердің түзілуі лецитин-холестеринді ацилтрансферазаның (LHAT) қатысуымен жүреді, ол лецитиннің i-күйінен холестеринге ацил қалдықтарының ауысуын катализдейді (10.31-сурет) LHAT бауырда пайда болады, қанда секрецияланады және липопротеиндерге қосылады. Әр түрлі липопротеиндердің эстерификация деңгейі айтарлықтай ерекшеленеді және LHAT (негізінен apo-AT, сонымен бірге CI) немесе осы ферментті тежейтін аполипопротеидтердің болуына байланысты (apo-AT болатын HDL-дегі ең белсенді LHAT). керек барлық белоктар olee 2/3. олеин және линол қышқылдардың күрделі эфирлері құрылған. қалыптастыру HDL қарағанда баяу қарқынмен жүреді сложноэфирные басқа липопротеидов ірі коли үздігі.
Сур. 10.31. LHAT әсерінен холестеринді эфирлердің түзілуі
LHAT HDL беткі қабатында локализацияланған және фосфолипидті монолайердегі холестеринді субстрат ретінде қолданады. Мұнда түзілген холестерин эфирлері толық гидрофобтығына байланысты аз сақталады
Метаболизм және липидтердің қызметі
фосфолипидті монолайзер және липопротеиннің липидті өзегіне батырылған. Сонымен қатар, фосфолипидті монолайерде холестеринге арналған орын босатылады, оны жасуша мембраналарынан немесе басқа липопротеидтерден холестеринмен толтыруға болады. Осылайша, HDL LHAT әрекеті нәтижесінде холестериндік тұзақ болып көрінеді.
Өт қышқылының синтезі
Бауырда холестериннің бір бөлігі өт қышқылдарына айналады. Галл қышқылдарын холан қышқылының туындысы ретінде қарастыруға болады (10.32-сурет).
Холан қышқылы организмде пайда болмайды. Гепатоциттерде холестерин тікелей ценодегихолдық және хол қышқылдарын - алғашқы өт қышқылдарын шығарады (10.33-сурет, 10.12-суретті қараңыз).
Холестерин биосинтезі
Холестерин биосинтезі эндоплазмалық ретикулумда жүреді. Молекуладағы барлық көміртек атомдарының көзі митохондриядан митохондриядан, майлы қышқылдар синтезінде пайда болған ацетил-SCoA болып табылады. Холестерин биосинтезі 18 ATP молекуласын және 13 NADPH молекуласын тұтынады.
Холестериннің пайда болуы 30-дан астам реакцияда жүреді, оларды бірнеше сатыға топтастыруға болады.
1. Мевальон қышқылының синтезі.
Алғашқы екі синтез реакциясы кетогенез реакцияларымен сәйкес келеді, бірақ 3-гидрокси-3-метилглютарил-ScoA синтезінен кейін фермент енеді гидроксиметил-глютарил-ScoA редуктазы (HMG-SCOA редуктаза), мевалон қышқылын түзеді.
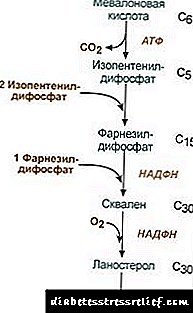 |
Холестеринді синтездеу реакциясы2. Изопентенил дифосфатының синтезі. Бұл кезеңде үш фосфаттың қалдықтары мевалон қышқылына қосылады, содан кейін ол декарбоксилденеді және дегидрленеді. 3. Изопентенил дифосфатының үш молекуласын біріктіргеннен кейін фарнессилфосфат синтезделеді. 4. Скваленнің синтезі екі фарнсилилфосфаттың қалдықтары байланған кезде пайда болады. 5. Күрделі реакциялардан кейін сызықты сквален ланостеролға ауысады. 6. Артық метил топтарын жою, молекуланы қалпына келтіру және изомеризациялау холестериннің пайда болуына әкеледі. Гидроксиметилглютарил-S-КоА редуктазасының белсенділігін реттеу3. Холестерин биосинтезінің жылдамдығы концентрацияға да байланысты нақты тасымалдаушы ақуызгидрофобты аралық синтез метаболиттерін байланыстыруды және тасымалдауды қамтамасыз етеді. Сіз өз пікіріңізді сұрай аласыз немесе қалдыра аласыз. Реттеудің негізгі мәні - мевалон қышқылының түзілу реакциясы. 1. Аллостерикалық реттеу. Холестерин, бауырда - және өт қышқылдары HMG-CoA редуктазасын тежейді. 2. HMG-CoA редуктазалық холестерин синтезінің репрессиясы. 3. Белсенді фосфорланбаған түрдегі HMG-CoA редуктазасының фосфорлану-депосфорлануымен реттеу. Глюкагон дезактивацияны тудырады, ал инсулин реакциялардың күрделі каскады арқылы активацияны тудырады. Осылайша, сіңіргіш және постабсорбциялық күйлердің өзгеруімен холестерин синтезінің жылдамдығы өзгереді. 4. Бауырдағы HMG-CoA редуктаза синтезінің жылдамдығы тәуліктік ауытқуларға ұшырайды: түн ортасында максимум және таңертең минимум. Холестерол эфирінің алмасуы Жасушаларда холестеринді этерификациялау әсер етілген кезде пайда болады ацил-КоА-холестерин-ацилтрансфераза (AHAT): Ацил-КоА + холестерин ® NS-CoA + ацилхолестерин Жасушаларда негізінен линолейхолестерол түзіледі. Эфирлер негізінен цитозол құрамында липидтер тамшыларының бөлігі ретінде кездеседі. Эфирлердің пайда болуы, бір жағынан, артық холестеринді мембранадан шығару механизмі, екінші жағынан, холестеринді жасушада сақтау тетігі ретінде қарастырылуы мүмкін. Қорларды жұмылдыру ферменттердің қатысуымен жүреді эфиразагидролиздеуші холестерол эфирлері: Ацилхолестерин + N2О ® Майлы қышқыл + холестерол Эфирлердің синтезі мен гидролизі әсіресе бүйрек үсті кортексінің жасушаларында белсенді болады. Қанда липопротеидтер эфирлердің пайда болуы қатысады лецитин-холестерин-ацилтрансфераза (LHAT), ацилді қалдықты лецитиннен холестеринге ауыстыруды катализдейді. LHAT бауырда пайда болады, қанға секрецияланады және липопротеидтерге қосылады. HDL-дегі ең белсенді LHAT, ол жер қабатында локализацияланған. Мұнда пайда болған холестеринді эфирлер гидрофобты және липидтер өзегіне батырылған. Фосфолипидті монолайерде холестерин үшін бос орын болады, оны жасуша мембраналарынан немесе басқа липопротеидтерден холестеринмен толтыруға болады. Осылайша, HDL LHAT әрекеті нәтижесінде холестериндік тұзақ болып көрінеді. Өт қышқылының синтезі Бауырда холестериннің бір бөлігі өт қышқылдарына айналады. Өт қышқылдары холан қышқылының туындылары ретінде қарастырылуы мүмкін. Холан қышқылы организмде пайда болмайды. Холестериннен гепатоциттерде бастапқы өт қышқылдары түзіледі - ценодеоксихикалық және холистикалық. Ішек флорасының ферменттерінің әсерінен ішекке өт шыққаннан кейін олардан екінші реттік қышқылдар түзіледі - литохолиялық және дезоксихологиялық. Олар ішектерден сіңіп кетеді, портал тамырының қаны бауырға, сосын өтке енеді. Өт құрамында негізінен конъюгацияланған өт қышқылдары, яғни олардың глицин немесе таурин бар қосылыстары бар. Өттегі өт қышқылдарының концентрациясы шамамен 1% құрайды. Өт қышқылдарының негізгі бөлігі қатысады гепатоэнтериялық айналым.Өт қышқылдарының аз бөлігі - күніне 0,5 г - нәжіспен шығарылады. Бұл бауырдағы жаңа өт қышқылдарының синтезі арқылы өтеледі, өт қышқылы қоры шамамен 10 күнде жаңартылады. Холестерин сонымен бірге ішек арқылы шығарылады. Ол ішектерге тамақпен және бауырдан өт құрамына енеді. Қанға сіңірілген холестерин құрамында өт пайда болатын фракция бар (эндогендік холестеринбауырда синтезделеді) және тағамнан алынған бөлшек (экзогендік холестерин) Холестеринді ұлпалардан шығару оның бауырдағы өт қышқылдарына дейін тотығуымен, содан кейін оларды нәжіспен (күніне шамамен 0,5 г) және өзгермеген холестеринді (нәжіспен) шығару жолымен жүреді. Тұрақты күйде: (Холестеринсоңы + Холестеринэкс) - (Холестеринэкранда + Өт қышқылдарыэкранда) = 0 Егер бұл тепе-теңдік бұзылса, тіндер мен қандағы холестерин концентрациясы өзгереді. Қандағы холестериннің жоғарылауы - гиперхолестеринемия. Бұл атеросклероз мен өт тас ауруының ықтималдығын арттырады. ЛИПИД АЛМАСУ РЕТІ Липидтер алмасуы орталық жүйке жүйесімен реттеледі. Ұзақ жағымсыз эмоционалды стресс, қанға катехоламиндердің шығарылуының жоғарылауы айтарлықтай салмақ жоғалтуға әкелуі мүмкін. Әрекет глюкагон липолитикалық жүйеде катехоламиндердің әсеріне ұқсас. Адреналин және норепинефрин тіндік липазаның белсенділігі мен май тініндегі липолиз жылдамдығын жоғарылатады, нәтижесінде қан плазмасындағы май қышқылдарының мөлшері артады. Инсулин адреналин мен глюкагонның липолизге және май қышқылдарының мобилизациясына кері әсерін тигізеді. Өсу гормоны аценилат циклазасын синтездейтін липолизді ынталандырады. Гипофиз гипофункциясы майдың ағзадағы тұндыруына әкеледі (гипофиздік семіздік). Тироксин, жыныстық гормондарлипидтер алмасуына да әсер етеді. Жануарлардағы жыныстық бездерді жою майды көп мөлшерде тұндырады. ЛИПИД МЕТАБОЛИКАЛЫҚ ДИСОРДОРЛАР Холестерин - бұл жануарлар организміне тән стероид. Оның адам ағзасындағы қалыптасуының негізгі орны - бауыр, онда холестериннің 50% синтезделеді, 15-20% -ы аш ішекте, қалғаны теріде, бүйрек үсті безі мен гонадтарда синтезделеді. Холестерин қорын қалыптастыру көздері және оны жұмсау тәсілдері 22.1 суретте келтірілген. Сур. 22.1. Денедегі холестериннің пайда болуы және таралуы. Адам ағзасындағы холестеринді (жалпы мөлшері шамамен 140 г) шартты түрде үш бассейнге бөлуге болады: 30 г) тез алмасады, ішек қабырғасының холестеринінен, қан плазмасынан, бауырдан және басқа паренхималық мүшелерден тұрады, жаңару 30 күнде жүреді (күніне 1 г), 50 г) баяу алмасатын басқа мүшелер мен тіндердің холестеринін, 60 г) жұлын мен мидың, дәнекер тіннің холестерині өте баяу алмасады, жаңарту жылдамдығы жылдар бойынша есептеледі. Холестерин синтезі жасушалардың цитозолында жүреді. Бұл адам ағзасындағы метаболизмнің ең ұзақ жолдарының бірі. Ол 3 сатыда жүреді: біріншісі мевалон қышқылының түзілуімен, екіншісі скалиннің пайда болуымен аяқталады (30 көміртек атомынан тұратын сызықтық көмірсутек құрылымы). Үшінші кезеңде скалалин ланостерол молекуласына айналады, содан кейін ланостеролды холестеринге айналдыратын 20 қатарлы реакциялар болады. Кейбір ұлпаларда холестериннің гидроксил тобы эфирлер түзеді. Реакция жасушаішілік AHAT ферментімен катализденеді (acylCoA: холестеринді ацилтрансфераза). Эфирлеу реакциясы HDL-де қанда пайда болады, онда LHAT ферменті (лецитин: холестеринді ацилтрансфераза) орналасқан. Холестеринді эфирлер - бұл оны қан арқылы немесе жасушаларға енгізу формасы. Қанда холестериннің шамамен 75% -ы эфирлер түрінде болады. Холестерин синтезі процестің негізгі ферментінің белсенділігі мен мөлшеріне әсер етеді - 3-гидрокси-3-метилглютарил-КоА редуктазы (HMG-CoA редуктазы). Бұған екі жолмен қол жеткізіледі: 1. HMG-CoA редуктазасының фосфорлануы / депосфорлануы. Инсулин HMG-CoA редуктазасының депфосфорлануын ынталандырады, осылайша оны белсенді күйге айналдырады. Сондықтан сіңіру кезеңінде холестерин синтезі жоғарылайды. Осы кезеңде ацетил-КоА синтездеуге арналған субстраттың қол жетімділігі артады. Глюкагон керісінше әсер етеді: ақуыз киназасы А арқылы белсенді емес күйге айналдырып, HMG-CoA редуктазасының фосфорлануын ынталандырады. Нәтижесінде постабсорбция кезеңінде және ораза кезінде холестерин синтезі тежеледі. 2. HMG-CoA редуктазының синтезін тежеу.Холестерин (метаболизм жолының соңғы өнімі) HMG-CoA редуктаза генінің транскрипция жылдамдығын төмендетеді, осылайша өзінің синтезін тежейді, өт қышқылдары да осындай әсер етеді. Қандағы холестеринді тасымалдау препараттың құрамында жүзеге асырылады. ТҚ ұлпаларда экзогендік холестеринді қамтамасыз етеді, оның ағзалар мен ағзалар арасындағы ағынын анықтайды. Экзогендік холестерин бауырға қалдық ХМ бөлігі ретінде жеткізіледі. Онда синтезделген эндогендік холестеринмен бірге ортақ қорды құрайды. Гепатоциттерде ТАГ мен холестерин VLDL-ге оралған және осы түрінде қанда шығарылады. LP-липазаның әсерінен ГЛГ-ны глицерол мен май қышқылдарына гидролиздей отырып, VLDLPs алдымен STLPs-ке, содан кейін 55% -ке дейін холестерин мен оның эфирлерінен тұратын LDLP-ге айналады. LDL - бұл холестериннің негізгі көлік формасы, ол арқылы ол тіндерге жеткізіледі (холестерин мен қандағы эфирлердің 70% -ы LDL құрамына кіреді). Қаннан алынған LDL бауырға (75% дейін) және олардың бетінде LDL рецепторлары бар басқа тіндерге енеді. Егер жасушаға түсетін холестерин мөлшері оның қажеттілігінен асып кетсе, онда LDL рецепторларының синтезі басылады, бұл қандағы холестерин ағынын азайтады. Жасушадағы бос холестерин концентрациясының төмендеуімен, керісінше, рецепторлардың синтезі іске қосылады. LDL рецепторларының синтезін реттеуге гормондар қатысады: инсулин, триодотиронон және жыныстық гормондар рецепторлардың түзілуін жоғарылатады, глюкокортикоидтер азаяды. «Холестериннің тасымалдануы» деп аталатын жерде, яғни. холестериннің бауырға оралуын қамтамасыз ететін жол, HDL басты рөл атқарады. Олар бауырда жетілмеген прекурсорлар түрінде синтезделеді, олар құрамында холестерин мен TAG болмайды. Қандағы HDL прекурсорлары холестеринмен қаныққандықтан, оны басқа сұйықтықтар мен жасуша мембраналарынан алады. Холестериннің HDL-ге ауысуы олардың бетінде орналасқан LHAT ферментін қамтиды. Бұл фермент май қышқылының қалдықтарын фосфатидилхолиннен (лецитин) холестеринге дейін қосады. Нәтижесінде холестеринді эфирдің гидрофобты молекуласы түзіледі, ол HDL ішінде жүреді. Осылайша, холестеринмен байытылған HDL мас емес HDL 3-ке айналады - жетілген және үлкенірек бөлшектер. HDL 3 VLDL және STD құрамындағы липопротеидтер арасында холестерол эфирлерін өткізетін белгілі бір ақуыздың қатысуымен холестеринді эфирлерді TAG-ға ауыстырады. Бұл жағдайда HDL 3 HDL2-ге айналады, оның мөлшері TAG мөлшерінің жиналуына байланысты артады. LLD-липазаның әсерінен VLDL және STDL негізінен холестеринді бауырға жеткізетін LDL-ге айналады. Холестериннің аз бөлігі HDL2 және HDL бауырына жеткізіледі. Өт қышқылдарының синтезі. Бауырда күніне 500-700 мг өт қышқылы холестериннен синтезделеді. Олардың түзілуіне гидроксилазалардың қатысуымен гидроксил топтарын енгізу және холестериннің бүйір тізбегінің ішінара тотығу реакциясы кіреді (22.2-сурет): Сур. 22.2. Өт қышқылдарының түзілу схемасы. Алғашқы синтез реакциясы - 7-а-гидроксиолестериннің түзілуі - реттеуші. Бұл реакцияны катализдейтін ферменттің қызметі жолдың соңғы өнімі, өт қышқылдары арқылы тежеледі. Реттеудің тағы бір тетігі - ферменттің фосфорлануы / депосфорлануы (7-а-гидроксилазаның фосфорланған формасы белсенді). Фермент мөлшерін өзгерту арқылы да реттеу мүмкін: холестерин 7-а-гидроксилаза генінің транскрипциясын тудырады, өт қышқылдары репрессияға ұшырайды. Қалқанша безінің гормондары 7-а-гидроксилаза синтезін, эстроген репрессиясын тудырады. Эстрогеннің өт қышқылдарының синтезіне әсері әйелдерде өт тасты ауруы ер адамдарға қарағанда 3-4 есе жиі кездесетінін түсіндіреді. Холестериннен пайда болған холликолды және ценодеоксихол қышқылдары «бастапқы өт қышқылдары» деп аталады. Бұл қышқылдардың негізгі бөлігі конъюгациядан өтеді - өт қышқылының карбоксил тобына глицин немесе таурин молекулалары қосылады. Конъюгация өт қышқылдарының белсенді формасын - CoA туындысын құрудан басталады, содан кейін таурин немесе глицин қосылады, нәтижесінде коньюгаттардың 4 нұсқасы пайда болады: таурохольдік және таурохенодезоколиялық, гликохолиялық және гликохенодезоксикалық қышқылдар. Олар бастапқы өт қышқылдарына қарағанда едәуір күшті эмульгаторлар. Глицинмен конъюгаттар тауринге қарағанда 3 есе көп түзіледі, өйткені организмдегі таурин мөлшері шектеулі. Ішекте бактериялық ферменттердің әсерінен бастапқы өт қышқылдарының аз мөлшерде қосылыстары екінші реттік қышқылдарға айналады. Холиктен түзілген және дезоксихолдан түзілген литохол қышқылы аз еритін және ішекте баяу сіңеді. Ішекке енетін өт қышқылдарының шамамен 95% -ы бауыр арқылы портал арқылы қайтып келеді, содан кейін қайтадан өтке секрецияланып, майлар эмульсиясында қолданылады. Өт қышқылдарының бұл жолы энтерофатиялық айналым деп аталады. Нәжіспен көбінесе қайталама өт қышқылдары алынады. Өт тас ауруы (холелитиаз) - бұл патологиялық процесс, оның негізі холестерин болып табылатын өт қабында тастар пайда болады. Холестеринді өтке шығару өт қышқылдары мен гидрофобты холестерин молекулаларын мицелярлық күйде ұстап тұратын пропорционалды шығарумен қатар жүруі керек. Өт қышқылдары мен холестериннің арақатынасының өзгеруіне әкелетін себептер: холестеринге бай тағам, жоғары калориялы тамақтану, өт қабындағы тоқырау, энтерофатиялық айналымның бұзылуы, өт қышқылдарының синтезінің бұзылуы, өт қабының инфекциясы. Холелитиазбен ауыратын науқастардың көпшілігінде холестерин синтезі жоғарылайды, одан өт қышқылдарының синтезі баяулайды, бұл өт ішіне бөлінген холестерин мен өт қышқылдарының санындағы диспропорцияға әкеледі. Нәтижесінде холестерин өт қабына түсіп, тұтқыр тұнба түзіп, біртіндеп қата бастайды. Кейде ол билирубинмен, ақуыздармен және кальций тұздарымен сіңдіріледі. Тастар тек холестериннен (холестерин тастарынан) немесе холестерин, билирубин, ақуыздар мен кальций қоспасынан тұруы мүмкін. Холестеринді тастар әдетте ақ, ал аралас тастар әртүрлі реңктерде қоңыр болады. Тас түзілуінің бастапқы кезеңінде ценодегихол қышқылы дәрі ретінде қолданыла алады. Өт қабында бір рет холестерин тастарын біртіндеп ерітеді, бірақ бұл бірнеше айға созылатын баяу процесс. Атеросклероз - бұл тамыр қабырғасының ішкі бетінде атерогендік бляшкалардың пайда болуымен сипатталатын патология. Мұндай патологияның дамуының негізгі себептерінің бірі - холестеринді тамақтан қабылдау, оның синтезделуі және организмнен шығуы арасындағы тепе-теңдіктің бұзылуы. Атеросклерозбен ауыратын науқастарда LDL және VLDL концентрациясы жоғарылаған. HDL концентрациясы мен атеросклероздың даму ықтималдығы арасында кері байланыс бар. Бұл LDL-нің ұлпалардағы холестеринді және тіндерден HDL тасымалдаушысы ретінде жұмыс істеу тұжырымдамасына сәйкес келеді. Атеросклероздың дамуының негізгі метаболикалық «алғы шарты» гиперхолестеринемия болып табылады. (қандағы жоғары холестерин). Гиперхолестеринемия дамиды: 1. холестеринді, көмірсулар мен майларды артық тұтыну салдарынан, 2. LDL немесе apoB-100 рецепторларының құрылымындағы тұқым қуалайтын ақаулардан тұратын генетикалық бейімділік, апоБ-100 синтезі немесе секрециясының жоғарылауы (қанның концентрациясы мен холестерин мен TAG жоғарылайтын отбасылық біріктірілген гиперлипидемия жағдайында). Атеросклероздың даму механизмдерінде препараттың модификациясы маңызды рөл атқарады. LDL құрамындағы липидтер мен ақуыздардың қалыпты құрылымындағы өзгерістер оларды ағзаға жат етеді, сондықтан фагоциттердің ұсталуы үшін қол жетімді. Дәрі-дәрмектің модификациясы бірнеше механизмдермен жүруі мүмкін: Қандағы глюкоза концентрациясы жоғарылағанда пайда болатын белоктардың гликозилденуі 2. липопротеидтердегі липидтердің өзгеруіне және апоБ-100 құрылымына әкелетін пероксидтің модификациясы, 3. LP-антиденесінің аутоиммунды комплекстерінің түзілуі (өзгертілген дәрілер аутоантиденелердің түзілуіне әкелуі мүмкін). Модификацияланған LDL макрофагтармен сіңіріледі. Бұл процесс сіңірілген холестериннің мөлшерімен реттелмейді, өйткені ол белгілі бір рецепторлар арқылы жасушаларға енеді, сондықтан макрофагтар холестеринге көп түсіп, субэндотелий кеңістігіне енетін «көбік жасушаларына» айналады. Бұл қан тамырлары қабырғасында липидті дақтардың немесе жолақтардың пайда болуына әкеледі. Бұл кезеңде тамырлы эндотелий өзінің құрылымын сақтай алады. Көбік жасушаларының санының өсуімен эндотелий зақымдалады. Зақым тромбоциттердің белсенділенуіне ықпал етеді. Нәтижесінде олар тромбоциттерді шығарады, тромбоциттердің агрегациясын ынталандырады, сонымен қатар тегіс бұлшықет жасушаларының көбеюін ынталандыратын тромбоциттерден алынған өсу факторларын шығара бастайды. Соңғысы медиальдан артериялық қабырғаның ішкі қабатына ауысады, осылайша бляшканың өсуіне ықпал етеді. Одан әрі бляшка талшықты тінмен өсіп, талшықты мембрананың астындағы жасушалар некротикалық, ал холестерин жасушааралық кеңістікке түседі. Дамудың соңғы кезеңдерінде бляшек кальций тұздарымен сіңдіріліп, өте тығыз болады. Бляшка аймағында қан ұйығыштары жиі пайда болады, бұл тамырдың люменін бітеп тастайды, бұл тиісті тін аймағында жедел қан айналымының бұзылуына және инфаркттың дамуына әкеледі. Холестерин синтезін реттеу - оның негізгі ферменті (HMG-CoA редуктаза) әртүрлі жолдармен жүзеге асырылады. HMG редуктазасының фосфорлануы / депосфорлануы. Инсулин / глюкагон коэффициентінің жоғарылауымен бұл фермент фосфорилатқа түсіп, белсенді күйге өтеді. Инсулиннің әрекеті 2 фермент арқылы жүзеге асырылады. HMG-CoA редуктазасы киназды фосфатаза, ол киназаны белсенді емес депосфорланған күйге айналдырады: Фосфотаза HMG-CoA редуктаза оны депосфорланған белсенді күйге айналдыру арқылы. Бұл реакциялардың нәтижесі HMG-CoA редуктазасының депфосфорланған белсенді формасының түзілуі болып табылады. Демек, сіңу кезеңінде холестерин жоғарылайды. Осы кезеңде холестерин - ацетил - КоА синтезі үшін бастапқы субстраттың қол жетімділігі артады (құрамында көмірсулар мен майлар бар тағамды жеу нәтижесінде, глюкоза мен май қышқылдарының ыдырауы кезінде ацетил Коа түзіледі). Постабсорбциялық жағдайда протеингеназа А арқылы глюкагон HMG - CoA - редуктазаның фосфорлануын ынталандырады, оны белсенді емес күйге айналдырады. Бұл әрекет глюкагонның HMG-CoA редуктазасының фосфорлануын және инактивациясын ынталандырады, осылайша HMG-CoA редуктазасын фосфорланған белсенді емес күйде ұстайды. Нәтижесінде постабсорбция кезеңінде және ораза кезінде холестерин синтезі эндогендік синтезге кедергі келтіреді. Егер тамақ құрамындағы холестерин мөлшері 2% -ға жеткізілсе, онда эндогенді холестерин синтезі күрт төмендеді. Бірақ холестерин синтезін толық тоқтату пайда болмайды. Тағамнан келетін холестериннің әсерінен холестерин биосинтезінің тежелу дәрежесі әр адамға байланысты өзгереді. Бұл холестериннің түзілу процестерінің жеке ерекшелігін көрсетеді. Холестерин синтезінің қарқындылығын төмендету арқылы оның қандағы концентрациясын төмендетуге болады. Егер бір жағынан холестеринді тамақпен қабылдау және оның ағзадағы синтезі мен екінші жағынан өт қышқылдары мен холестериннің шығуы арасындағы тепе-теңдік бұзылса, тканьдер мен қандағы холестерин концентрациясы өзгереді. Ең ауыр зардаптар қандағы холестерин концентрациясының жоғарылауымен байланысты (гиперхолестеринемия), ал атеросклероз мен холелитиаздың даму ықтималдығы артады. Отбасылық гиперхолестеринемия (HCS) - бұл форма әлдеқайда жиі кездеседі - 200 адамға шамамен 1 науқас. HCS-тегі мұрагерлік ақау LDL-нің жасушаларға сіңуін бұзады, сондықтан LDL катаболизмі жылдамдығының төмендеуі болып табылады. Нәтижесінде қандағы LDL концентрациясы, сондай-ақ холестерин жоғарылайды, өйткені LDL-де оның мөлшері көп. Сондықтан, HCS-мен холестеринді тіндерге, атап айтқанда теріге (ксантомалар), артерия қабырғаларына тұндыру тән. HMG-CoA редуктазының синтезін тежеу Холестериннің метаболизм жолының соңғы өнімі. Ол HMG-CoA редуктаза генінің транскрипция жылдамдығын төмендетеді, осылайша өзінің синтезін тежейді. Бауыр холестериннен өт қышқылдарын белсенді түрде синтездейді, сондықтан өт қышқылдары HMG-CoA редуктаза генінің белсенділігін тежейді. HMG-CoA редуктаза шамамен 3 синтезден кейін болғандықтан, бұл холестерин ферментінің синтезін тежеу тиімді реттеу болып табылады.
|